
|
|
||
|
A-
CHRONOLOGIE DES DECOUVERTES SUR LE SOMMEIL |
||
|
||||||||||||
|
Qui
parle de rêve, parle
aussi de sommeil et donc bien sûr du cerveau. Le
rêve est
un ensemble de mécanismes et de conditions très
complexes. Pour l'étudier, il nous faudra parler tout
d'abord de
l'aspect général de celui-ci dans le sommeil. L'année
suivante, en 1950, le neurobiologiste lyonnais Michel
Jouvet découvre la
phase de rêve, «le
sommeil paradoxal». Alors que tous les muscles sont inertes,
le
cerveau est en pleine activité. |
||
|
B-
LA DIVISION DU SOMMEIL EN TROIS PHASES |
||
|
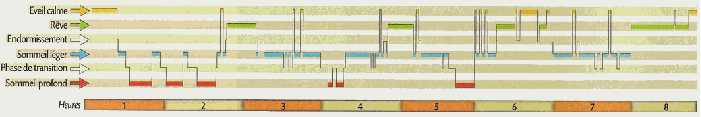
Le
sommeil qui jusqu'à
maintenant n'était considéré que comme
une seule
phase, est en fait un ensemble comprenant trois phases. Le
sommeil paradoxal apparaît lorsque le système
d'éveil ainsi que celui de l'endormissement sont totalement
désactivés. Le sommeil paradoxal est une phase du
sommeil
pendant laquelle l'activité électrique corticale
est
similaire à celle de l'éveil. On
peut aussi remarquer l'apparition des mouvements oculaires
rapides ainsi que la disparition du tonus musculaire. C'est une atonie
posturale. On retrouve pendant ces périodes, des moments
d'érections de vingt-cinq minutes ainsi que l'activation de
l'activité ponto geniculo occipital (PGO). Sourd, aveugle et
paralysé, le dormeur devient alors très
vulnérable. C'est pour cela qu'il ne peut s'endormir que
s'il se
sent en totale sécurité. Cet
ensemble de phases est très bien structuré et est
entrecoupé par des périodes très
courtes de
réveil. Ces « micro
éveil » peuvent
se produire plusieurs fois par nuits. |
||

|
Après
ce bref descriptif des différentes phases du sommeil, nous
allons nous concentrer sur le sommeil paradoxal, période
pendant
laquelle intervient les rêves, sujet de notre TPE. |
||
|
C-
LE FONCTIONNEMENT DU SOMMEIL |
||
|
Nous
avons dit que le sommeil
paradoxal est une phase pendant laquelle l'animal est très
vulnérable car il est totalement impuissant. L'atonie
posturale est une paralysie totale du corps dans une position
momentané.Elle est déclenchée par un
petit groupe
de neurones de taille moyenne situés dans le locus coeruleus
alpha et sa partie interne. Ces neurones sont sensibles à
l'acétylcholine. Cette substance provoque une atonie
généralisée accompagnée
soit du sommeil
paradoxal soit de l'éveil (état de cataplexie). A
la base de cette atonie posturale, c'est une
activité ponto géniculo occipital (PGO). Cette
activité est située dans la formation
réticulée pontique. Les voies qu'empreinte cette
activité mène au cortex
cérébral par
l'intermédiaire parfois de relais thalamiques. Ce
mécanisme est appelé le frein moteur. Il est
divisé en trois étages. Le premier est
l'étage des
commandes et les deux autres sont des étages
d'exécution. Dans
l'immense majorité des rêves, voire dans tous les
cas, le
rêve est visuel. Les rêves sonores sont moins
fréquent (45% des rêves). On y éprouve
des
sensations tactiles et corporelles dans 35% des cas. Le goût
et
l'odorat sont largement sous représentés avec 1%
des cas.
Beaucoup de nos rêves sont des cauchemars ou des semi
cauchemars. |
||
|
Exemple
de la vue d'un banquier : |
||

|
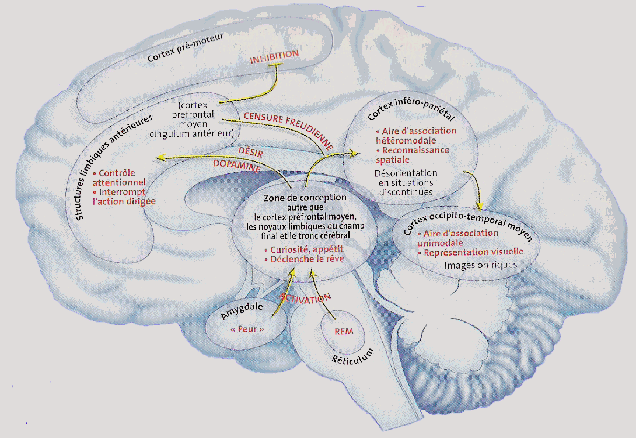
Que
se passe-t-il lorsque l'on ferme les yeux et qu'il n'y a aucune
information visuelle ? Les aires primaires visuelles sont
désactivées et pourtant, l'imagerie interne
persiste. En
fait, toutes ces régions du cerveau produisent leurs propres
images que la volonté peut susciter. « Le
cerveau produit en permanence ses propres
images si bien que certains psychologues estiment qu'en fait nous
hallucinons la réalité plus que nous la
voyons » explique Sophie Schwartz. Les
régions
cérébrales du traitement de l'image
réalisent ce
que les spécialistes appellent des interférences
en
créant une sorte d'image idéale qui n'est pas du
tout
celle perçue par la rétine. La simple impression
de
relief est une construction du
cerveau. « Face à
un paysage, on ne perçoit pas une image faite de points,
lignes
et couleurs, mais des montagnes, des arbres... » Durant
le sommeil paradoxal, ces régions très actives
pourraient
perdre leur cohérence ou se mettre à fonctionner
indépendamment du reste du cerveau, comme lors de certaines
pathologies mentales. L'absence d'informations visuelles
empêcherait les inférences oniriques
d'être
réajustées. En outre, le lobe
préfrontal
étant peu actif pendant cette phase de sommeil,
l'élimination des images absurdes ou incongrues ne se ferait
pas
correctement. Le rêveur serait alors un spectateur de ses
propres
hallucinations, un naïf croyant aux effets spéciaux
de son
cerveau. |
||
|
D-
LE RÔLE DU RÊVE, DU SOMMEIL PARADOXAL |
||
|
On
peut tout d'abord
préciser que seulement 10% des spécialistes
contestent la
thèse de M.Jouvet selon laquelle le sommeil
paradoxal est
l'équivalent physiologique du rêve. |
||



